电脑系统除了微软的还有其他的么-电脑系统除了gjo还有
1.电脑上出现了一个空文件夹,打不开也删不掉
2.求计量软件stata百度云资源下载,我的电脑是XP系统的。拜托了
3.木霉表达系统
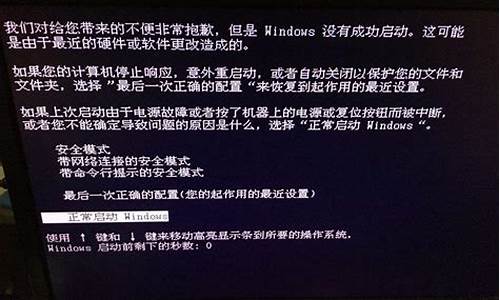
4.电脑中了木马或病毒,无法启动瑞星和360,想要重装,但没光盘,怎么办?
电脑上出现了一个空文件夹,打不开也删不掉

1.
先确认是否为系统文件,系统隐藏文件夹可能会删除再出现。
2.
很有可能是软件安装后自带的资料夹。
3.
安装360,右键该文件夹,选择强力删除后粉碎文件夹。
4.
将文件夹名称修改掉,右键选择删除或清理删除。
求计量软件stata百度云资源下载,我的电脑是XP系统的。拜托了
点击下载:链接:提取码:1717?
链接:提取码:1717?
链接:提取码:1717?
有的下载了软件,可是愁于安装需要激活,人家也不是免费给你用的,接着要去百度,stata序列号,stata激活码,stata破解工具之类的,啊啊啊,我的头脑都要爆炸了。怎么下载一个软件这么麻烦啊,到底有没有能够让我用的软件?
真的没有吗?世上无难事,只怕有心人。小编告诉你,你不是一个人在战斗,你还有一大群的小伙伴和你一样在抗战。还有我。
自己学习的时候老师分享的资料,分享给你,包括软件和详细图文安装教程。
Stata是一套提供其使用者数据分析、数据管理以及绘制专业图表的完整及整合性统计软件。它拥有很多功能,包含线性混合模型、均衡重复反复及多项式普罗比模式。用Stata绘制的统计图形相当精美。
木霉表达系统
长期以来,丝状真菌在食品、发酵等工业领域中有着广泛应用,尤其在工业酶制剂的生产中占据了核心地位。工业酶制剂的国际市场份额每年高达数十亿美元,并且在不断增长,其中近40%的产量来自丝状真菌。因此,丝状真菌很快成为颇具吸引力的生产异源重组蛋白的表达系统。
大多数情况下,自然界中的野生菌株生产目标蛋白的水平很低,不能直接用于工业生产。为了得到高产量的目标蛋白,需对野生菌株进行遗传改造。随着分子遗传技术和真菌基因转移系统的发展,利用基因工程方法对丝状真菌进行遗传改造已经成为可能。
T.reesei作为表达系统具有许多优点:第一,它本身能够释放大量的纤维素酶,具有强启动子序列,可以在一定程度上提高外源蛋白的产量;第二,它能够有效地释放和分泌蛋白,大大降低了下游工艺的难度和成本;第三,它的工业应用已经相对成熟,以它为受体菌生产各种蛋白质的工艺会相对简洁;第四,它不仅具有适于蛋白生产的诸多优点,同时对人没有毒性,在产酶条件下也不产生真菌毒素和抗生素。近年来的实践表明,经过基因工程手段改造的T.reesei重组菌株有很高的安全级别,应用潜力巨大。
在蛋白质表达的研究中,影响蛋白产量的因素包括启动子强度及启动子拷贝数、信号肽等。另外,内源基因的高效表达可能会影响外源蛋白的产量。
高效表达的基因,其启动子必定具有较高的转录起始效率。选择强启动子构建外源基因表达系统,是提高外源蛋白质表达量的一种重要策略。另外,启动子与受体细胞之间的合理匹配是外源基因在宿主细胞中表达的关键。同源启动子有利于受体细胞调控因子的识别,减少甲基化,促进外源基因整合到染色体上,从而提高转化效率和外源基因的表达效率(Hirano et al.,1999)。在T.reesei表达外源蛋白时,cbh1,cbh2,eg1和eg2等主要内源基因在机体内相对高效的表达势必会影响外源基因的表达,其分泌的胞外蛋白质中,纤维二糖水解酶I的含量占总分泌蛋白的50%,高效表达的基因,其启动子必定具有较高的转录起始效率,纤维二糖水解酶I的启动子(cbh1)被认为是T.reesei中最强的启动子,用其构建表达载体来生产同源或是异源蛋白是非常理想的。王肖燕(2012)通过构建T.reesei主要内源纤维素酶基因缺失菌株时发现:cbh1缺失菌株Δcbh1外切葡聚糖酶I的量与出发菌株Δtku70相比明显下降,而外切葡聚糖酶Ⅱ的分泌量有所提高;cbh2缺失菌株Δcbh2::pyrG+外切葡聚糖酶I的量与出发菌株Δtku70相比没有太大变化,外切葡聚糖酶Ⅱ的分泌量有所下降;cbh1/cbh2双缺失菌株Δcbh1Δcbh2外切葡聚糖酶I和外切葡聚糖酶II的量与出发菌株Δtku70相比均明显下降。目前已经利用cbh1强启动子构建了木霉高效表达载体(吕丹丹,2012;汪天虹等,2003),研究证明,利用cbh1强启动子和终止子构建表达载体,均能提高内源和外源基因的表达(Margolles-Clark et al.,1996 f;母敬郁等,2006;龙昊,2006;金欣等,2011;王冰冰等,2011;董欣睿等,2012)。Joutsjoki(1993)构建了两种一步基因置换载体。一种载体含有Hormoconis resinae葡萄糖淀粉酶P的基因组基因(gamP),另一种含有相应的cDNA,都在T.reesei cbh1启动子控制下表达。这些载体经转化后置换T.reesei的cbh1基因。在这两种载体中,cbh1启动子都精确地连接到gamP蛋白质编码区上游,指导T.reesei分泌有活力的葡萄糖淀粉酶P(GAMP)。研究结果表明,含有gamP cDNA的最优转化体能分泌大约700mg/L有活力的GAMP,是在H.resinae中产量的20倍(Joutsjoki et al.,1993)。Karlsson等(2001)将egl4基因置于强启动子序列cbh1之后,构建了Pcbh1-egl4-Tcbh1表达盒,并转化入T.reesei进行表达,对EGI和EGIV活力进行比较,结果显示,EGIV的cmC,Avicel,HEC 活力都明显低于EGI,在内切酶活力中的作用较小。Miettinen-Oinonen等(2002)把比活力较高的egl2基因插入cbh1启动子和终止子之间,并且定向整合到染色体cbh2基因片段,使之失活,构建了一个强内切酶活力而外切酶活力较低的T.reesei 重组菌株。Miettinen-Oinonen等(2005)将cbh1和cbh2基因片段定向整合到egl1和egl2上,构建了一个强外切酶活力而内切酶活力较低的新型T.reesei,来满足不同的产业需求。Wang等(2004)构建了新型表达盒Pcbh1-egl3-Tcbh1,并且定向整合到染色体cbh1上,使之失活,分析重组菌的CMC活力,是原始菌株的两倍,而FPA活力则显著下降。Haakana等(2004)从Melanocarpusal-bomyces真菌中克隆到三种不同的纤维素酶基因,分别为编码20kDa和50kDa内切葡聚糖酶蛋白的cel45A,cel7A基因,以及编码50kDa外切葡聚糖酶蛋白的cel7B基因,并且在T.reesei中进行了表达,结果显示他们能够在中性pH下高效工作,较酸性p H的纤维素酶应用更加广泛。R ib e iro等(2010)研究了T.reesei的外切葡聚糖酶基因cbh1和内切葡聚糖酶基因eg1在棉阿舒囊霉Ashbya gossypii中的表达,同时与在酿酒酵母S.cerevisiae中的表达情况进行了对比,发现egI基因的表达水平比要cbh1高,两者均存在过糖基化现象,但比在酿酒酵母中程度低(Jianping et al.,2011)。
研究发现,经过多拷贝策略改造的cbh1 启动子,在顺式功能区4 拷贝以内,随着cbh1启动子拷贝数的升高,报告基因的表达水平呈上升趋势。使用多拷贝的启动子作为外源蛋白表达的启动子,能提高启动子的活性,增加目的蛋白的表达量。为构建人源促红细胞生成素在T.reesei中的高效表达载体,刘宣(2007)利用T.reesei cbh1强启动子基础上构建的4拷贝人工启动子,cbh1终止子和CBHI信号肽序列构建了分泌型的人源EPO表达载体pE。将pE质粒与携带潮霉素磷酸转移酶基因(hph)的pAN7-1 质粒分别共转化T.reesei蛋白酶缺陷型菌株RUT C-30 U4和在RUT C-30 U4基础上进一步经过初步糖基化改造,已经转入人源N-乙酰氨基葡萄糖转移酶I基因的工程菌株T108,在含有150mg/L潮霉素的基本培养基平板上各筛选得到78个和150个潮霉素抗性转化子,经PCR验证,epo基因已经被整合到转化子染色体上。RT-PCR,SDS-PAGE和Western Blot杂交分析结果表明epo基因在T.reesei中已经被转录和翻译,表明重组人源EPO蛋白在T.reesei中成功表达。SDS-PAGE和Western Blot杂交分析结果显示带有N-乙酰氨基葡萄糖转移酶I基因的转化子所表达的蛋白的糖基化程度有所提高。
N末端信号肽的作用是引导分泌蛋白进入分泌途径。在外源蛋白的生产中,通常把来自分泌良好的内源蛋白的信号肽序列与欲构建的重组蛋白融合,以引导其分泌。当外源蛋白本身就是胞外蛋白时,其内部的信号序列可以用来指导分泌。烟曲霉(Aspergillus fu-migatus)AfMp1p是一个由284个氨基酸组成的半乳甘露聚糖蛋白,属于典型的 GPI-CWP。苏建臣等(2013)将AfMp1p的细胞壁定位GPI 信号肽和烟曲霉几丁质酶AfChiB1的N-端信号肽分别与绿色荧光蛋白(Green Fluorescent Protein,GFP)的C末端和N末端融合并转化T.reesei。荧光观察结合Western blot的结果表明,在平台期中期和后期,带有GPI信号的GFP融合蛋白定位于细胞壁。这说明烟曲霉来源的GPI信号可被T.reesei识别,从而用于介导外源蛋白在T.reesei细胞壁中的定位表达。
随着蛋白质组的发展,研究者开始利用融合蛋白标签,可以获得高纯度和形态结构明确的重组蛋白。融合蛋白上的蛋白质标签与包被在固相基质上的特异性的配基集合,可以达到对目的蛋白的分离纯化的目的。融合蛋白标签根据分子量大小可以分为蛋白质分子(或结构域及其衍生物)和多肽片段两类。大多数情况下,由于多肽标签较小,对融合蛋白质结构影响小,不需要从融合蛋白中切除,更为常用。融和蛋白标签与基质的作用类型主要包括酶与底物,细菌受体与血清蛋白,聚组氨酸与金属离子,抗原与抗体等。蛋白分泌过程及蛋白分泌的动力学研究的资料由Pakula等(2000)通过体内标签实验获得。通过研究发现,CBHI纤维二糖水解酶的平均合成时间为4min,平均分泌时间为11min,在S.cerevisiae和其他丝状真菌中,它的分泌时间要更长些。这说明T.reesei具有良好的蛋白分泌能力的原因不仅分泌速度快,更主要是因为它具有高效的、大容量的分泌途径。Pa-kula等(2000)获得CBHI的各种不同pI形式的形成过程。最先形成的是最小的酸性pI形式,折叠抑制剂(DTT)处理的细胞中只有这种形式。内质网形成后开始出现更多酸性pI形式,去糖基化实验也证明这些酸性pI形式来自CBHI聚糖结构的修饰。蛋白的生产效率也随菌株的生长时期而变(Pakula et al.,2005),在菌株低生长速度下,CBHI和其他分泌蛋白的合成与分泌效率最高达到0.03g·L-1·h-1。在低生长速率下蛋白分泌速率成为限制CBHI产量的主要因素,同时内质网中涉及折叠蛋白的编码基因pdi1和bip1的转录水平就显得比较高,这说明细胞需要依据生长速率调整内质网中折叠效率。
结语
本章详细地论述了木霉的蛋白分泌途径、糖基化过程及其相关蛋白基因。但是仍然还有很多具体的细节仍在推测阶段,有些重要阶段分泌我们知道得较少,很多重要的蛋白酶的功能仍在探究中,目前较低的木霉转化率和表达外源蛋白的不稳定性仍是制约木霉大规模应用的主要因素。但是,木霉的表达系统可以克服在原核表达系统中无法进行特定翻译后修饰的缺陷,而且比酿酒酵母具有更高的分泌外源蛋白的能力,所以成为生产具有生物活性的真核生物蛋白的重要途径。目前在木霉中特别是T.reesei中已发展多种分子系统,采用不同的策略用于同源和异源蛋白的生产,为了提高蛋白产量,使用强表达和有效分泌的纤维素酶CBHI基因的不同部分,已被证明是一种非常有效的方法,这都得益于cbh1启动子很强,而且其位点也易于表达。通过完善生产策略和技术,将会不断拓宽木霉生产重组蛋白的范围。随着现代分子生物学研究的进展及基因工程在木霉中的应用,木霉的蛋白分泌与糖基化途径将被完全揭开神秘的面纱,我们相信,木霉将成为人类利用木霉生产多种酶和药用产品的重要工具。
电脑中了木马或病毒,无法启动瑞星和360,想要重装,但没光盘,怎么办?
可以下载wsyscheck,我教你,运行后你看到有些系统进程是粉色的,这些一般是有病毒插入了一些文件起到开机自动启动和保护的功能,你点击一下,看的一些很陌生的dll或exe文件,你点直接删除,但是这你要十分了解你的系统,应为有些可能不是病毒文件,是杀毒软件或是你的程序的文件,所以小心点删,你可以根据文件厂商来判断,再点击内核检查,看的ssdt和fsd检查 ,把里面的代码异常写了yes的点恢复代码函数,在系统模块里有些不认识的点击直接删除,服务管理,安全检查,里的不认识的,名字很乱(比如很多数字,很多无规律的,明显是乱打的,或是数字和英文结合的文件),都删除,然后在文件管理里,查找当天中病毒时产生的文件,一个一个看,很可疑的都删除,然后点击注册表管理,里面你看到个框框,点下箭头,你会看到有很多的注册表路径,一个一个点,把里面的可以的文件启动项删掉,还是按照我前面说的判别可疑的方法。
然后下载sreng2,要改名字,不然病毒不会让他运行,打开后,如果会有什么提示什么被修改,就按照他说的点右键恢复,在系统修复里,点击里面的高级修复,默认的就可以了,点击修复,然后点击下面的修复安全模式,apihook检查,就可以了,
然后进入安全模式查杀就行了
如果还不懂,在线mm我
声明:本站所有文章资源内容,如无特殊说明或标注,均为采集网络资源。如若本站内容侵犯了原著者的合法权益,可联系本站删除。